

作者:维尔克斯 时间:2026-7-7 14:06:30
本文介绍一种结构光照明显微镜(含MLA微透镜阵列超分辨成像或结构光照明显微镜MLA微透镜阵列或结构光照明MLA微透镜阵列)的模拟实现方法,能够实现三维超分辨率成像(MLA微透镜阵列成像),横向分辨率为145 nm,轴向分辨率为350 nm,采集速度最高可达100 Hz。通过采用光学图像处理操作而非数字图像处理操作,我们无需捕获、存储和组合多个相机曝光,从而将数据采集速率比其他超分辨率显微镜提高了10到100倍,并能够实时采集和显示超分辨率图像。低激发强度允许对数百个二维切片进行成像,结合物理和计算切片,可实现与转盘共聚焦显微镜相当的深度穿透能力。
现有的超分辨率荧光显微镜为了获得亚衍射极限的样品信息,往往需要牺牲采集速度。我们报道了一种结构光照明显微镜的模拟实现方法,能够实现三维超分辨率成像(MLA微透镜阵列成像),横向分辨率为145 nm,轴向分辨率为350 nm,采集速度最高可达100 Hz。通过采用光学图像处理操作而非数字图像处理操作,我们无需捕获、存储和组合多个相机曝光,从而将数据采集速率比其他超分辨率显微镜提高了10到100倍,并能够实时采集和显示超分辨率图像。低激发强度允许对数百个二维切片进行成像,结合物理和计算切片,可实现与转盘共聚焦显微镜相当的深度穿透能力。我们通过对精细、快速移动的结构(包括人肺成纤维细胞中马达驱动的细胞器以及发育中的斑马鱼胚胎内流动血细胞的细胞骨架)进行成像,展示了我们系统的能力。
现代荧光显微镜结合了对比度、分子特异性、速度和生物相容性,能够实现活细胞过程的可视化。然而,衍射将宽场荧光显微镜的分辨率限制在横向约250 nm、轴向500–750 nm,而且在实际中要达到这种“衍射受限”的性能本身就颇具挑战。超分辨率成像技术[1]克服了这一难题,但在现有实现方案中,速度、成像时长和视场都受到严重影响,特别是与传统显微镜相比。例如,单分子成像[2,3]或受激发射损耗(STED)显微镜[4]能够在细胞区域实现<100 nm的空间分辨率,但成像速度被限制在约0.01–1 Hz(更快的成像需以牺牲视场和图像质量为代价)[5]。无论是单分子成像还是STED,目前都难以用于长时间序列的体积成像,原因在于采集速度低以及存在光毒性照明强度(峰值强度10⁴–10⁷ W/cm²,平均功率数十毫瓦)。
与单分子成像和STED相比,线性结构光照明显微镜[6,7](SIM)提供的分辨率提升较为适中(比衍射极限高√2倍,解卷积后高两倍),但所需的照明强度低10³–10⁶倍,采集速率快得多(二维可达11 Hz,三维可达0.2 Hz)[8,9],并且能通过计算去除离焦光。这些优势使得在整个细胞体积内的数百个三维时间点上实现光学层切超分辨率成像成为可能。通过采用稀疏激发点阵并结合发射光路中的针孔以及适当的图像处理(补充说明)来物理性地抑制离焦光,SIM的穿透深度可以进一步提高[10–13],从而实现距离盖玻片表面约50 µm深处的实时超分辨率成像[13](称为“多焦点SIM”或MSIM)。无论具体实现方式如何,以往的工作都需要用约10–100个激发图案照明样品,每个图案采集一次相机曝光,然后将所得图像进行数字合成以生成一张二维超分辨率图像。这种每层需要捕获并合成多张原始图像的需求,从根本上限制了SIM相对于传统显微镜的速度。
我们报道了一种模拟实现的结构光照明显微镜技术,能够在完全不牺牲速度、光毒性或视场的前提下,将荧光显微镜的空间分辨率提高一倍。通过消除采集和数字合成多幅相机曝光图像的需求,我们的方法去除了SIM相较于传统荧光显微镜的唯一缺点,实现了超分辨率图像的实时采集与显示。我们的技术能够以与线扫描或转盘共聚焦显微镜相当或更快的速率进行多色体积成像,从而可以双色观察亚线粒体细节——这些细节原本会被衍射或运动模糊所掩盖。我们进一步通过以比先前报道快15倍的体积成像速率,对相互作用的蛋白分布进行无创超分辨率成像,凸显了本方法相对于现有SIM实现方案的优势。最后,我们通过以37 Hz的速率实时可视化流动血细胞内的细胞骨架细节,并以100 Hz的速率捕捉内质网的毫秒尺度重塑与生长过程,展示了前所未有的超分辨率成像速度。
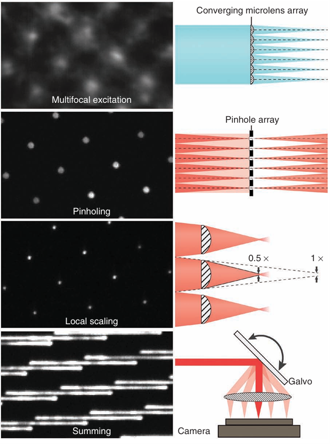
图1 | 实现即时结构光照明的关键步骤
一个会聚MLA微透镜阵列超分辨成像用于产生多焦点激发。激发样品后,通过与MLA微透镜阵列超分辨成像相匹配的针孔阵列抑制离焦荧光。借助第二个相匹配的结构光照明显微镜MLA微透镜阵列,每个针孔后的荧光发射实现两倍的局部收缩。振镜用于对多焦点激发进行光栅扫描并叠加多焦点发射,从而在每次相机曝光期间生成一幅超分辨率图像(为清晰起见,仅显示了部分振镜扫描)。左侧为每个步骤对应的原始数据,右侧为示意图(另见补充图1)。
结果
用于即时超分辨率的模拟图像处理
我们方法的关键实现思路在于:在我们之前描述的MSIM技术中,数字组合的每一个步骤都可以通过光学方式以模拟、即时的方式进行(补充说明)。MSIM的数据采集和处理在概念上可分为多个步骤:(i) 对样品进行稀疏的多焦点照明;(ii) 用相机记录所产生的荧光图像;(iii) 数字针孔处理以抑制离焦发射;(iv) 对每个针孔后的发射进行数字两倍局部收缩(不改变整体图像尺寸);(v) 在多焦点激发图案的不同位置重复步骤(i–iv),直至整个视场被完全照明;(vi) 将所得图像进行数字叠加,生成分辨率提高√2倍的超分辨率图像;(vii) 解卷积以恢复完整的两倍分辨率提升。在这里,我们完全通过硬件实现步骤(i–vi)(图1),使用结构光照明显微镜MLA微透镜阵列产生多焦点激发图案,使用匹配的针孔阵列抑制离焦发射,使用匹配的结构光照明MLA微透镜阵列对每个针孔后的发射进行局部收缩,并使用振镜平移激发图案并在图像采集和相机叠加之前,对由这三个阵列产生的荧光发射进行叠加。
我们的即时结构光照明显微镜(“instant SIM”;补充图1-6)能够在单次相机曝光中直接捕获具有√2倍分辨率提升的光学切片图像,并在随后的解卷积步骤后产生分辨率提高一倍的图像。与MSIM类似,使用instant SIM对固定的人骨肉瘤(U2OS)细胞中Alexa Fluor 488标记的微管进行成像,结果显示微管表观宽度为150 nm,是宽场分辨率的两倍,使我们能够分辨中心间距为110 nm的微管(补充图7)。我们在100 nm荧光珠上验证了这一结果(145 ± 14 nm,平均值±标准差,n = 10),同时也获得了SIM所实现的轴向分辨率约两倍的提升(356 ± 37 nm)(表1和补充图8、9)。
快速且非侵入性的3D超分辨率成像(MLA微透镜阵列成像)
为了展示instant SIM在长时间序列全细胞体积成像方面的能力,我们对转化人肺成纤维细胞中的多种动态蛋白分布进行了成像。例如,我们构建了TFAM(一种线粒体DNA转录因子,同时也负责将线粒体DNA包装成核样体[14])的GFP嵌合体,并将其与Tom20(线粒体外膜转位酶复合物的成员,功能是选择性地将细胞质中的蛋白质转运到线粒体膜间隙)标记的mCherry[15](Tom20-mCherry)同时成像(图2a和补充视频1)。instant SIM的速度使我们能够在1.2秒内采集完一个双色、3 µm厚的体积数据(比以往对同等厚度样品进行的多色体积SIM快约15倍)[16],即使是以每秒0.5–1 µm速率快速迁移的线粒体也能清晰分辨(图2b)。我们的分辨率足够高,可以分辨出大多数线粒体内部的空洞(在Tom20通道中明显可见,补充图10)。TFAM核样体(其斑点大小处于我们的分辨率极限,与在固定细胞上进行的其他超分辨率实验结果一致[17])被包裹在线粒体空洞内(图2b),这一观察结果在样品的轴向视图中得到了证实(图2c)。据我们所知,STED是唯一被用来揭示线粒体内膜空间缺乏Tom20的其他超分辨率技术[18,19],但这些实验是在固定细胞上进行的,推测是由于STED采集速度慢或存在光毒性。
表1 | 不同显微镜下测得的100 nm亚衍射珠粒的表观宽度
|
|
即时SIM,宽场模式 |
即时SIM原始图像 |
即时SIM+解卷积 |
SDC原始图像 |
SDC+解卷积 |
LSC原始图像 |
LSC+解卷积 |
|
NA |
1.45 |
1.45 |
1.45 |
1.49 |
1.49 |
1.46 |
1.46 |
|
横向半高全宽(nm) |
281±11 |
213±26 |
145±14 |
260±11 |
188±14 |
369±30 |
250±38 |
|
轴向半高全宽(nm) |
680±30 |
511±24 |
356±37 |
633±36 |
429±20 |
661±31 |
401±14 |
Decon,解卷积;NA,数值孔径;SDC,转盘共聚焦;LSC,线扫描共聚焦;FWHM,半高全宽。“宽场模式”测量是在移除了结构光照明MLA微透镜阵列和针孔的即时SIM(instant SIM)上进行的。尽管数值孔径较低,但即时SIM中的珠粒看起来最清晰。数据来自10个珠粒,表示为平均值±标准差。“原始”值被用作本文中对数据集进行解卷积时所采用的理论高斯点扩散函数(PSF)的参数。
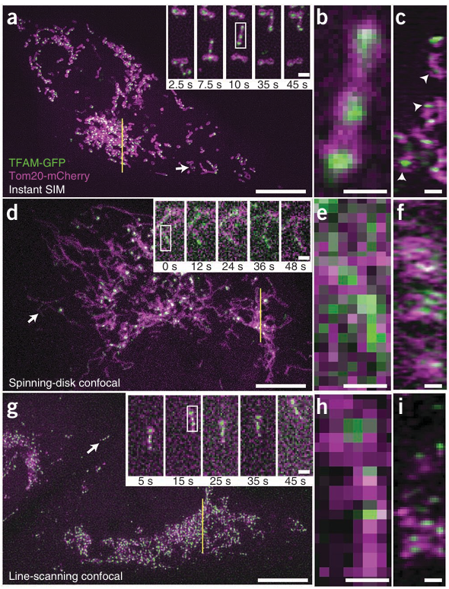
图2 | 即时SIM可实现高速双色超分辨率成像
(a–i) 对表达TFAM-GFP(绿色)和Tom20-mCherry(品红色)的活体MRL-TR转化人肺成纤维细胞,分别使用即时SIM (a–c)、转盘共聚焦 (d–f) 和快速线扫描共聚焦显微镜 (g–i) 进行成像。图中展示了3 µm厚体积的最大强度投影(xy),其中白色箭头所指的子区域在指定时间点以更高放大倍数显示(插图)。在g图的线扫描共聚焦插图中,图像相对于低倍视图旋转了90°。比例尺:10 µm(主图)和1 µm(插图)。b、e、h图显示了由a、d、g图插图中白色矩形所标示的线粒体更高放大倍数图像。比例尺:0.5 µm。c、f、i图显示了约270 nm厚切片的轴向视图(zy),切片位置对应于a、d、g图中的黄线。白色箭头指示了在即时SIM中观察到被Tom20包裹的TFAM。比例尺:1 µm(另见补充视频1–3和补充图8–12)。
从功能和概念上讲,即时SIM(instant SIM)很像一台分辨率更高的转盘共聚焦显微镜,因此我们将我们的方法与这种成熟且广受欢迎的技术进行了比较。对于同等厚度的双色体积,我们的转盘系统比即时SIM慢约10倍(补充视频2),并且横向和轴向分辨率均低于即时SIM(图2d–f,表1以及补充图8、9)。无论是否进行解卷积(图2e),或未进行解卷积(补充图11),转盘的分辨率都不足以观察线粒体内部的空洞(补充图12),并且使核样体变得模糊,以至于它们通常呈现为连续的而非离散的实体(图2e、f)。我们还与一种快速线扫描共聚焦显微镜进行了比较,该显微镜的体积采集速率与即时SIM相当(图2g–i和补充视频3)。与转盘共聚焦一样,线扫描共聚焦中线粒体空洞也变得模糊不清(补充图12),无论是否进行解卷积(图2h)或未进行解卷积(补充图11)。此外,与即时SIM相比,其轴向分辨率的损失足以使TFAM被Tom20包裹这一现象完全被衍射所掩盖(图2i,表1以及补充图8、9)。
即时SIM所采用的低激发强度(约5–50 W/cm²)使我们能够对足够明亮的样品进行数十个时间点的体积成像,而不会出现明显的光漂白或光损伤迹象(如膜回缩或起泡)。例如,我们可视化了丰富的H-Ras和Rab8A GTPases(生长因子信号级联和细胞内运输的关键因子[20])在60–100个成像体积中的动态变化(补充视频4和5)。即时SIM揭示了管状内吞体的形成和运动,以及基底膜附近高度移动的小囊泡(补充视频5)。鉴于近期对过氧化物酶体与线粒体相互作用的关注[21],我们对GFP标记的过氧化物酶体与Tom20-mCherry进行了活细胞同时成像(补充视频6)。值得注意的是,我们的时空分辨率还使我们能够可视化过氧化物酶体与线粒体球体的快速相互作用(补充视频7)。最后,我们使用GFP标记的内质网-过氧化物酶体护送蛋白(GFP-SEC16B),并对其与mCherry标记的过氧化物酶体的相互作用进行了成像(补充视频8),观察到了这两个伙伴之间的“亲吻并逃离”相互作用,这可能代表了从内质网到过氧化物酶体的物质转移[22](补充视频9)。在所有情况下,我们的采集速率比其他SIM实现方案[9,16]至少快15倍。
超越视频帧率的超分辨率成像
为了充分利用即时SIM的速度,我们在数百个时间点上以100 Hz的频率在单个二维平面内记录了内质网的动态变化(图3a和补充视频10)。我们选择内质网是因为其高度可塑性和快速运动[23]。我们的帧率使得我们能够轻松观察和量化单个内质网小管的快速生长(约3.5 µm/秒)和重塑(图3b)。我们还观察到了新生内质网小管的形成和生长,这些事件发生在140毫秒以内(图3c),而在任何其他SIM实现方案中,即使是以已报道的最快二维帧率[8]进行成像,这些事件也会变得模糊。我们注意到,尽管我们的成像面积大了40倍,但我们的成像速率仍比其他以类似有效分辨率记录内质网动态的方法[24]快50倍。
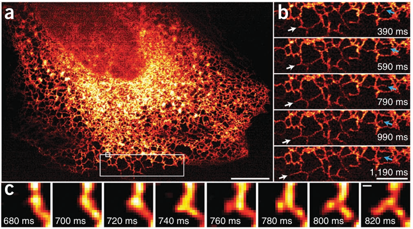
图3 | 即时SIM以100 Hz揭示内质网动态
(a) 200个时间点序列中的第一帧图像,显示MRL-TR转化人肺成纤维细胞内GFP-Sec61A标记的内质网。数据采集于盖玻片表面。比例尺,10 µm。(b) 图a中大白矩形框区域的高倍放大视图。白色箭头标示一个内质网小管的生长;蓝色箭头标示一个内质网小管的重塑。比例尺,5 µm。(c) 图a中小白矩形框区域的高倍放大视图,显示在140毫秒内形成的新小管。比例尺,200 nm(另见补充视频10)。
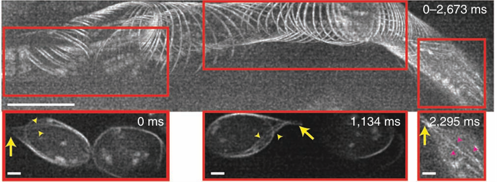
图4 | 以37 Hz帧率进行亚细胞分辨率的血管血流成像
上图:100帧二维图像(跨度2,673毫秒)的最大强度投影,显示位于3日龄斑马鱼胚胎头部血管内(深度20 µm)血细胞中的GFP标记微管。运动方向为从左向右。清晰的细胞边界表明无运动模糊。比例尺,10 µm。
下图:对应上图红色方框区域在指定时间点的空间局部放大图。黄色箭头:同一细胞末端的“尾巴”结构;黄色箭头标示:延伸到尾巴中的微管;品红色箭头标示:细胞内部的微管。比例尺,2 µm(另见补充视频11)。
即时SIM的一个优势在于,它结合了计算光学切片和物理光学切片,能够在比其他SIM实现方案[13]深约10倍的样品深度处进行超分辨率成像。我们利用这一特性,对距离盖玻片表面20–25 µm处、表达GFP转基因标记微管的3日龄斑马鱼胚胎头部血管内的血流进行了成像。我们的成像帧率为37 Hz,足以“定格”单个血细胞,揭示其亚细胞细节,尽管它们运动迅速(25 µm/秒,图4和补充视频11)。一些血细胞显示出由多个微管组成的“尾巴”(图4),此外还有横跨细胞体的细胞内微管。在某些情况下,微管束分散开来,揭示出很可能是单个微管的结构,这一点从其稀疏的标记可以推断。我们推测带有尾巴的细胞可能是中性粒细胞,因为这些尾巴在外观上类似于“吊索”结构,这种结构有助于中性粒细胞在高剪切力存在下滚动[25],尽管这些细胞的身份尚待确认。
讨论
与以往提供相当分辨率、光学切片能力和穿透深度的结构光照明显微镜[13]相比,即时SIM的速度快100倍。若计入处理时间,加速因子超过10,000。由于其近乎即时的采集与显示能力,即时SIM的成像速率超越了最先进的商用共聚焦显微镜,同时具有更好的分辨率。与STED和单分子成像技术相比,在相当的视场下,即时SIM快数个数量级。尽管如此,其时空分辨率、光效率和穿透深度仍有进一步提升的空间。
我们的三维采集速度受到样品台惯性的限制,但光学重聚焦策略[26]可以规避这一问题,并有望实现更高的体积成像帧率。我们的光学装置让人联想到一种扫场共聚焦系统[27],仅多了一个额外的光学元件(发射光路中的结构光照明MLA微透镜阵列)。这意味着商用硬件可能很容易被改装以提供即时SIM能力,但发射光路中相应数量较多的光学元件会降低信号水平,从而降低可实现的最大采集速度。我们注意到,即时SIM所需的多焦点激发、针孔滤波和局部收缩操作可以在转盘共聚焦几何结构中用更少的发射光学元件来实现,从而相应地提高速度和信号水平。使用多焦点激发必然意味着由于针孔间的串扰,其光学切片能力低于使用单个聚焦光束的激发。实际上,在我们选择的针孔间距下,即时SIM提供的切片能力与转盘共聚焦显微镜相当,但劣于点扫描或线扫描共聚焦显微镜(补充图13)。如果针孔间距更大,或者使用双光子激发,切片强度和穿透深度将得到改善。最后,如果使荧光发射强度非线性地依赖于照明强度(“非线性SIM”),即使在大视场和亚秒帧率下,空间分辨率也可以进一步提高[28,29]。因此,将光稳定、可逆切换的荧光蛋白[24,31]与即时SIM光学设计相结合,或许能够在100纳米以下的分辨率范围内实现视频帧率的超分辨率成像。
文章来源:A. G. York, P. Chandris, D. D. Nogare, J. Head, P. Wawrusin, R. S. Fischer, A. Chitnis, and H. Shroff, “Instant super-resolution imaging in live cells and embryos via analog image processing,” Nature Methods, vol. 10, no. 11, pp. 1122–1126, Nov. 2013, doi: 10.1038/nmeth.2687.



